3.1.A.1 – Enzyme Structure & Function in Biological Regulation
🧬 What Are Enzymes?
- Enzymes are proteins that act as biological catalysts — they speed up chemical reactions in cells without being used up.
- They work by lowering the activation energy, meaning reactions happen faster and more efficiently.
⚙️ Why Is Their Structure Important? 
- Enzymes have a specific 3D shape, especially at the active site (where the substrate binds).
- This structure is essential to their function. If it changes (like due to heat or pH), the enzyme may stop working (denaturation).
- The active site must match the substrate — like a lock and key (or more accurately, an induced fit that molds to the substrate).
💡 Key Idea:
🧩 The shape of the enzyme determines what reaction it catalyzes. If the shape is wrong, it can’t bind the substrate, and the reaction won’t happen properly.
🔁 Enzymes Regulate Processes by:
- Controlling reaction speed – reactions only happen when needed.
- Ensuring specificity – only target molecules are affected.
- Being reusable – the same enzyme can catalyze many reactions.
🧠 Summary:
- Enzymes are structured proteins that lower activation energy to speed up reactions.
- Their shape = function, and they play a key role in regulating almost all processes in living systems.
3.1.A.2 – Enzyme Specificity & Enzyme-Substrate Complex
🧩 How Do Enzymes Recognize Their Substrate?
For a chemical reaction to happen with an enzyme, two main things must match:
- Shape of the substrate must fit the enzyme’s active site
- Charge compatibility must also match (positive-negative attraction, etc.)
Think of it like a magnet puzzle – not only does the shape need to fit, but the charges must line up too.
🧪 The Enzyme-Substrate Complex
When the enzyme binds to its substrate, it forms an enzyme-substrate complex.
- This is a temporary structure the enzyme holds the substrate in the right orientation to allow the reaction to happen fast and efficiently.
- After the reaction, the product is released, and the enzyme is unchanged and reusable.
🔐 Models That Explain This: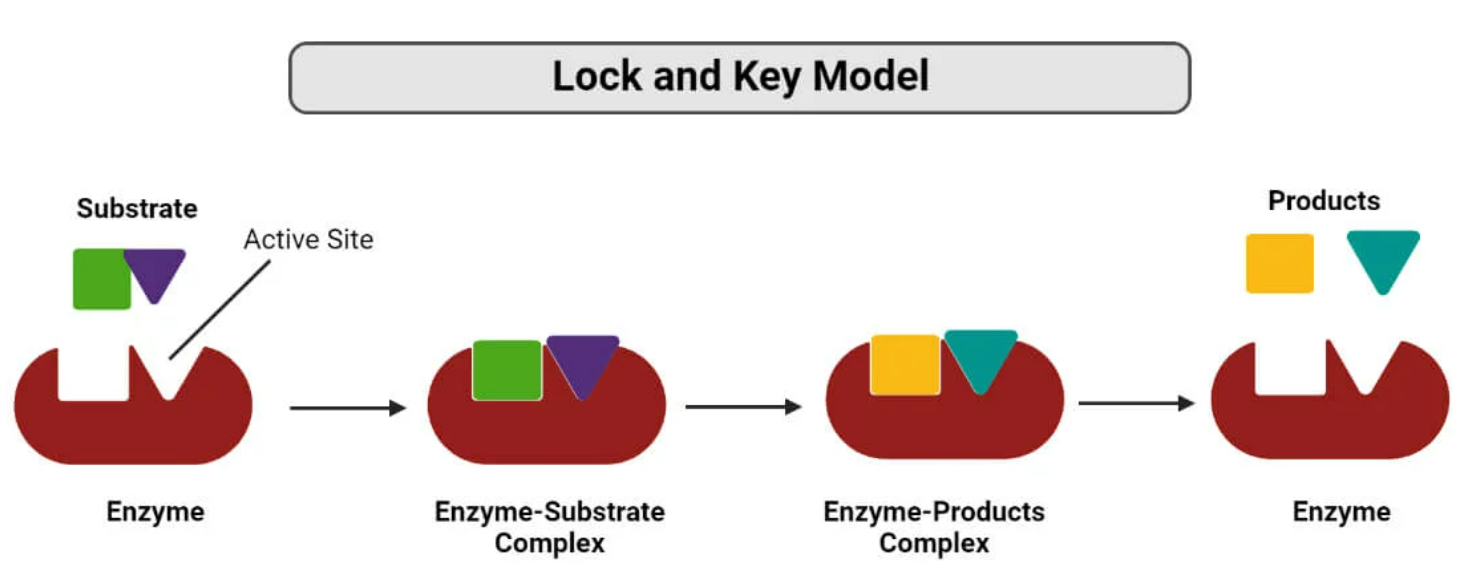
Lock-and-Key Model
- Substrate fits into the active site perfectly (like a key in a lock).
- Shows specificity but is a bit rigid.
Induced Fit Model (more accepted)
- The enzyme molds slightly around the substrate to get a tighter fit.
- Explains how enzymes can be flexible while still being specific.
🧠 Summary:
- Shape + charge compatibility between substrate and enzyme = reaction success
- Temporary enzyme-substrate complex allows fast, efficient reactions
- Induced fit model best explains enzyme flexibility and specificity